Kontaktinformation
Projektledare: fr.o.m. 2016-07-01 Frauke Ecke (tidigare Birger Hörnfeldt), Sveriges lantbruksuniversitet
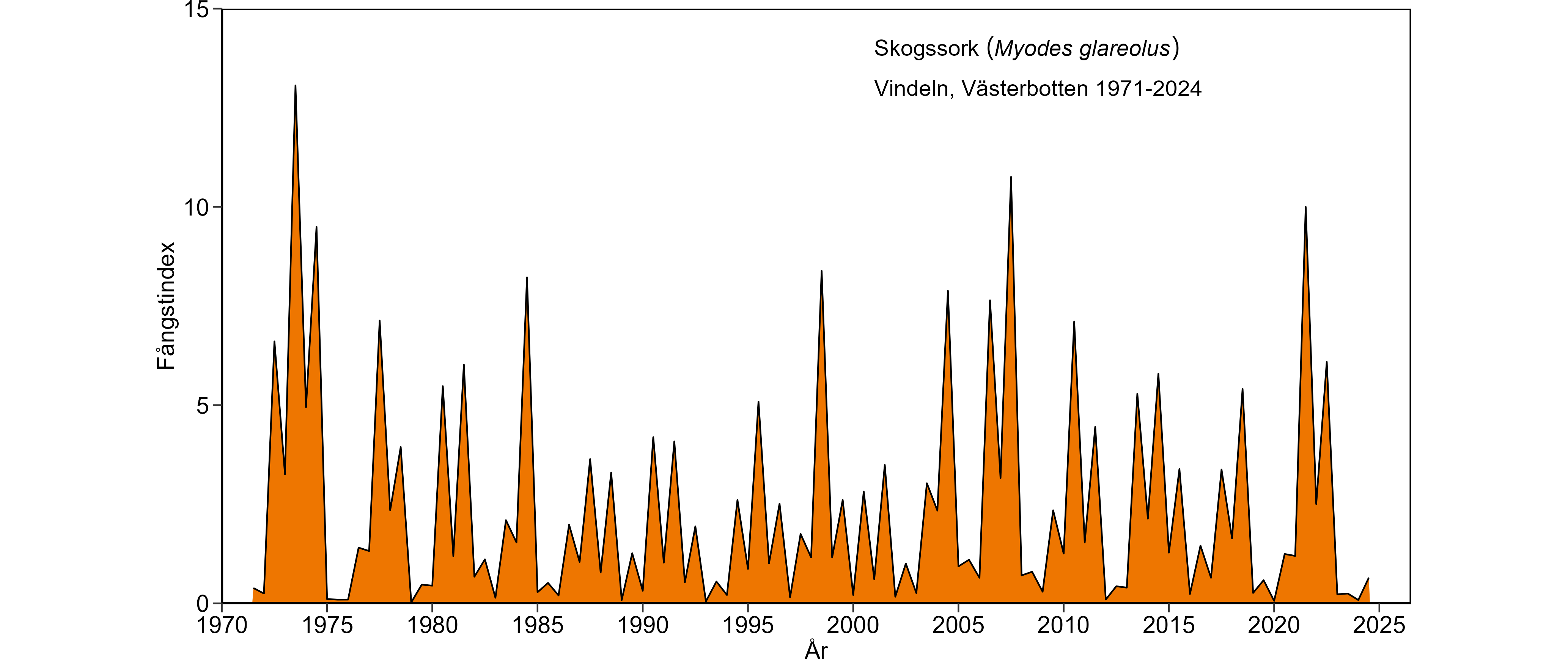
En ny sorkcykel inleddes våren 2020 med mycket låga sorktätheter. Under sommaren 2020 byggdes dock sorkpopulationen successivt upp. Den höga överlevnaden av skogssorkar under vintern 2020/21 resulterade hösten 2021 i de tredje högsta tätheterna för arten sedan miljöövervakningen startade 1971. Under 2022 var sorktätheten fortfarande hög, vilket resulterade i en dubbeltopp hösten 2021 och hösten 2022. Tätheterna under våren 2023 visade på en kraftig nedgång som också höll i sig under hösten.
Läget är något svårbedömt. Den kraftiga nedgången under 2023 följdes antingen åt av ett förstadium till en uppgångsfas eller så representerar 2024 en lågfas. Det senare scenariot innebär i så fall att cykeln som började 2020 är 5-årig. För en 5-årig cykel talar att uppgången under 2024 var mycket liten. Våren 2025 bör inledas med en märkbar uppgång av sorkpopulationen; dock från låga tätheter.
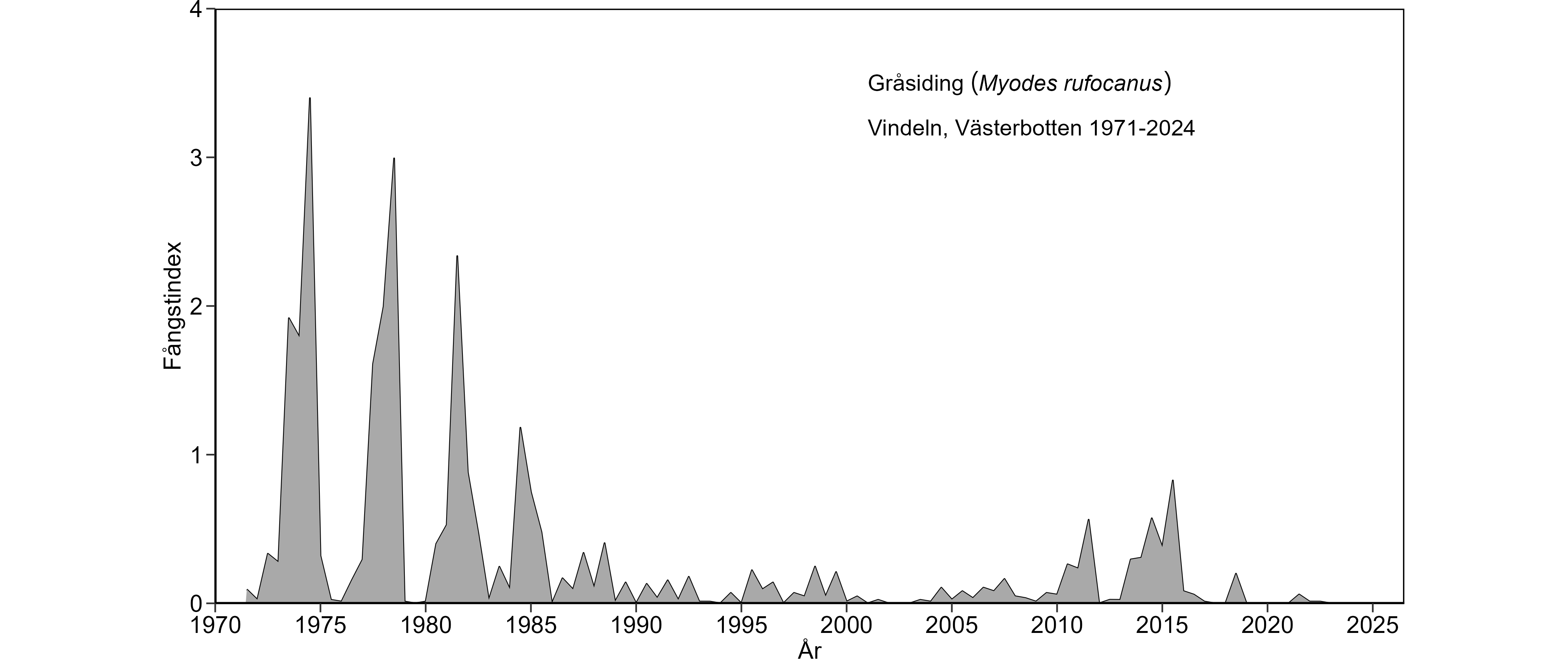
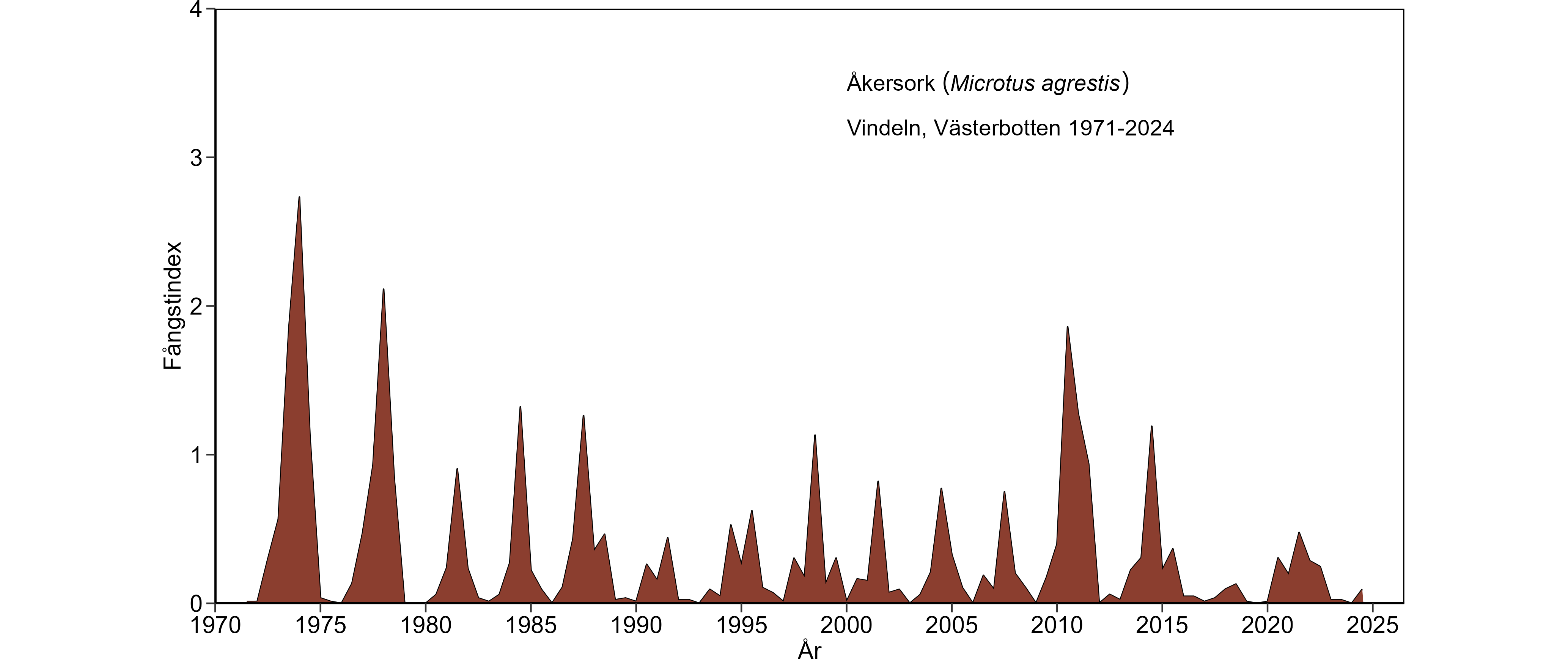
Gråsidingbeståndet i Västerbotten visade, vid sidan av 3-4-års cyklerna, en mycket tydlig nedåtgående långtidstrend följd av fortsatt låga tätheter under 1970-, 1980-, 1990- och 2000-talet. Nedgången var uppenbar både för vår- och hösttätheter. Även skogssork (tidigare: ängssork) och åkersork visade en minskad beståndsstorlek, men minskningen hos skogssork var inte lika dramatisk som för gråsidingen, medan utvecklingen för åkersorken på senare år ger anledning till oro. För skogssork och särskilt åkersork var främst vårtätheterna lägre under
1980-,1990-, 2000- och 2010-talen än under 1970-talet.
Mätresultaten från det första decenniet på 2000-talet förstärker bilden av att gråsidingens nedgång stått i en klass för sig. Täthetsindex i slutet av 2000-talet visade på en beståndsstorlek som var ungefär 1% av vad den var som mest i början av 1970-talet vid motsvarande fasläge av sorkcyklerna. Motsvarande värde för åkersorken var mindre än 10%. Sorkcyklerna, 2009-12 och 2013-15, kännetecknades av en svag återhämtning för gråsiding, så att läget då började se ut som i mitten på 1980-talet. Denna återhämtning har dock kommit av sig under senaste cyklarna. Åkersorken visade en stark återhämtning under sorkcykeln 2009-12, som däremot inte fortsatt under de senaste tre sorkcyklerna (2013-15, 2016-19 och 2020-pågående). Vintern 2018/19 var en nedgångsfas för skogssorken och avslutade denna 4-åriga cykel, dvs. 2016-2019. Tätheterna för åkersorken under den pågående cykeln var de lägsta sedan början av 1990-talet. Jämfört med sorkcykeln 2013-15 stannade gråsidingens återhämtning upp under cykeln 2016-19 och läget har inte förbättrats under pågående cykel.
Under perioden med minskande/låga sorkbestånd var skogssorkens minskning minst uttalad; exempelvis visade höststammens storlek relativt höga toppar såväl 1998, 2004, 2007 som 2010, väl i nivå med hur det såg ut i slutet av 1970-talet. Dessutom visade arten den starkaste uppgångsfasen hittills under 2006 med avseende på den beståndstäthet som nåddes på hösten, och som t o m var högre än efter uppgångsfasen 1972. Trots att det starka höstbeståndet 2006 följdes av en ovanligt kraftig vinternedgång för fasläget, var vår- respektive höststammen under 2007 de starkaste sedan början av 1970-talet.
En slående långtidsförändring, vid sidan av de minskade tätheterna, är det ökade säsongsmässiga inslaget i beståndsvariationerna. Både gråsiding och åkersork visade en högre andel vinternedgångar under 1980-, 1990-, 2000- och 2010-talen, vilket framgår av kurvornas ökade inslag av "sågtandsmönster" då jämfört med under 1970-talet. Skogssorkens beståndsförändringar har under hela tidsperioden karaktäriserats av mycket regelbundna vinternedgångar som orsakat kurvans "sågtandsmönster". Å andra sidan har skogssorkens vinternedgångar tenderat att vara starkare tidigare i cyklerna fr. o. m. 1980-talet än under 1970-talet. Sammantaget tyder detta på försämrade övervintringsförhållanden.
Varför har sorkarna minskat? Orsaken till gråsidingens och de andra arternas minskning är okänd förutom för gråsidingen. Det kan inte uteslutas att minskningen följt ett naturligt mönster, men det verkar troligare att minskningen indikerar någon form av miljöstörning. De försämrade övervintringsförhållandena tyder på att åtminstone en del av beståndsnedgången bör kunna förklaras av någon form av störning/förändring under vintern eller av någon störning som fått effekt just då. Sannolikt har de mildare vintrarna på senare år haft en negativ effekt, bl a genom att göra det skyddande snötäcket mer instabilt och kortvarigt och försämra dess kvalitet ur skyddssynpunkt.
En minskad förekomst och ökad fragmentering av viktiga biotoper för gråsidingens fortplantning, överlevnad och beståndsuppbyggnad till följd av avverkning inom skogsbruket verkar en drivande eller åtminstone bidragande faktor till gråsidingens särklassiga minskning (Magnusson et al. 2015b). Forskning har kopplat minskningen till sådan biotopförstöring på landskapsnivå. Arbetet har hittills legat till grund för tre doktorsavhandlingar. I den senaste avhandlingen visade Magnus Magnusson att gråsidingens förekomst på provytenivå, d.v.s. i landskapet, påverkas positivt av förekomsten av blockfält i det omgivande landskapet och hur väl blockfälten genom sammanbindande skog ”sitter ihop” med provytorna.
Gråsidingen som är en nordlig art, med utbredning ner längs den Skandinaviska fjällkedjan och därifrån ner i skogslandet, lever i Västerbotten i kanten av sitt utbredningsområde. I denna "utsatta position" kan man anta att gråsidingen är känsligare än övriga arter för miljöstörningar och därför reagerar snabbare och starkare på dessa.
Även om stammen för skogssork har återhämtat sig sedan mitten av 2000-talet måste man vara öppen för att det lika snart kan ske en återgång till minskande/låga sorkbestånd om vi får tillbaka de senaste årens mildare vintrar med sämre snöförhållanden. Försämrade vinterförhållanden förmodas slå extra hårt mot åkersork som föredrar öppna miljöer där risken för isbarksbildning under milda vintrar är stor. De låga tätheterna för åkersorken under de senaste cyklarna (2016-19, 2020-pågående) kan vara ett resultat av just sådana förhållanden. Om den vanligaste artern, skogssork, kommer att drabbas lika hårt som gråsidingen och på senare år även åkersork, så skulle det vara ett mycket hårt slag mot den biologiska mångfalden av ugglor, rovfåglar och rovdäggdjur.
Kemiska analyser av lever och njure från gråsidingar insamlade från en lokal, Ekträskkludden, i Västerbotten under 1980-97 och lagrade i Miljöprovbanken slutfördes under 1998 i ett pilotprojekt inom ramen för den Nationella miljöövervakningens miljögiftsprogram; i samarbete med Miljöprovbanken och Miljöforskningslaboratoriet vid SLU i Umeå (Hörnfeldt et al. opublicerat). Resultaten visade främst på en förändrad/störd mineralämnesbalans under den tidsperiod som var möjlig att undersöka retrospektivt med hjälp av materialet från Miljöprovbanken. Bland annat minskade kalciumhalterna i både lever och njure, medan kalium/kalcium- och fosfor/zink-kvoterna ökade och svavel/molybden-kvoten minskade.
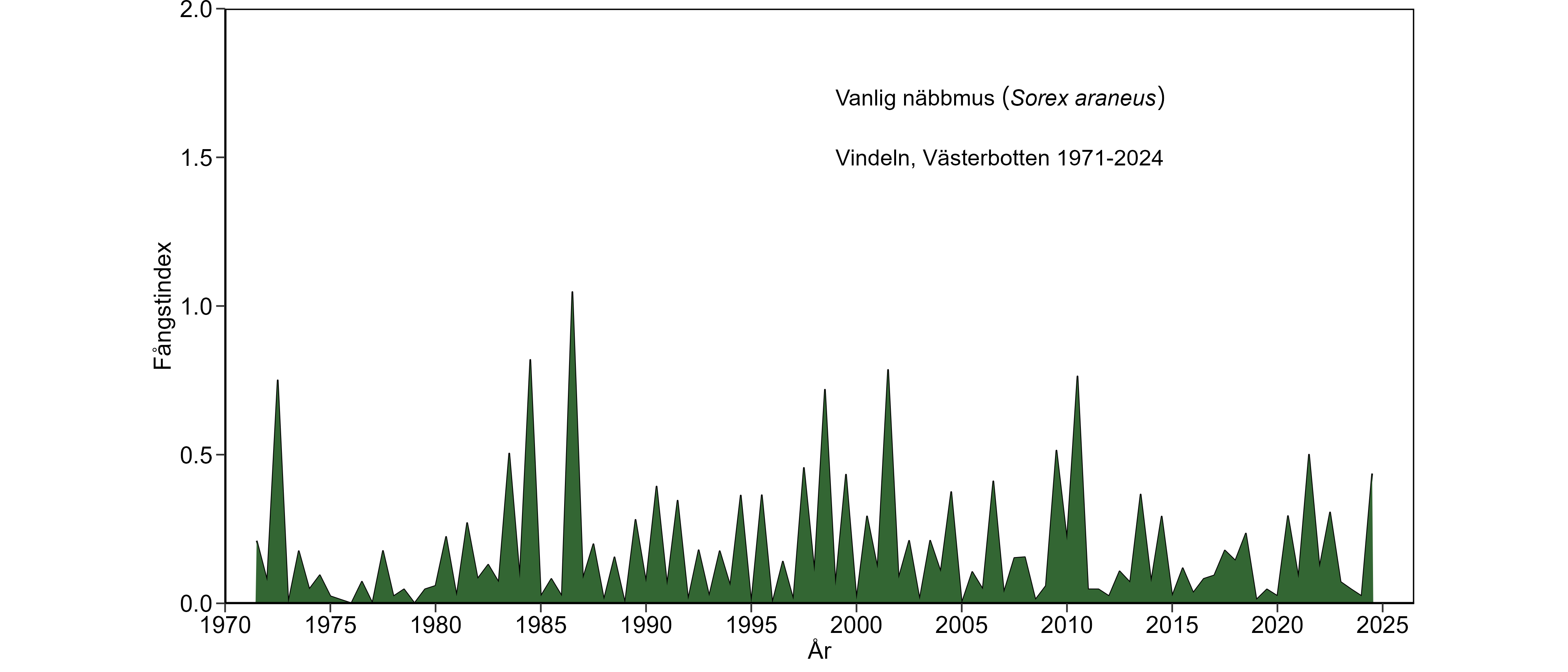
Upptäckten av den förändrade mineralämnesbalansen hos gråsiding är intressant, men orsaken är okänd. Det är även okänt om nedgången i gråsidingbeståndet är orsaksmässigt kopplad till den störda mineralämnesbalansen. Material från insamlade skogssorkar och åkersorkar i Miljöprovbanken har ännu inte analyserats för att klarlägga om liknande störningar av mineralämnesbalansen även förekommer hos dessa vanligare och för den biologiska mångfalden av rovdjur, därmed viktigare arter. En nyligen publicerad studie om förändringar av element och organiska ämnen hos skogssork i de olika miljöövervakningsområdena ger dock förutsättningen för att studera potentiella rubbningar i ämnesbalansen även hos skogssork..
En sammanfattande beskrivning och analys av minskningen i sorkbestånden och en närmare presentation av olika hypoteser om möjliga orsaker till minskningen har tidigare gjorts; här kan du läsa mer om minskningen och hypoteserna.
En analys av konsekvensen av minskande sorkbestånd för de rovdjur som lever av sorkar har även gjorts, med minskande bestånd av pärluggla som exempel; här kan du läsa mer om hur rovdjuren kan påverkas.
Christensen, P., Ecke, F., Sandström, P., Nilsson, M. and Hörnfeldt, B. 2008. Can landscape properties predict occurrence of grey-sided voles? Pop. Ecol. 50: 169-179 (DOI 10.1007/s10144-008-0077-5). (pdf)
Christensen, P. and Hörnfeldt, B. 2003. Long-term decline of vole populations in northern Sweden: a test of the destructive sampling hypothesis. J. Mammal. 84: 1292-1299.
Christensen, P. & Hörnfeldt, B. 2006. Habitat preference of Clethrionomys rufocanus in boreal Sweden. Landscape Ecology 21: 185-194.
Cornulier, T., Yoccoz, N.G., Bretagnolle, V., Brommer, J.E., Butet, A., Ecke, F., Elston, D.A., Framstad, E., Henttonen, H., Hörnfeldt, B., Huitu, O., Imholt, C., Ims, R.A., Jacob, J., Jędrzejewska, B., Millon, A., Petty, S.J., Pietiäinen, H., Tkadlec, E., Zub, K., Lambin, X. 2013. Europe-Wide Dampening of Population Cycles in Keystone Herbivores. Science 340: 63-66.
Ecke, F. & Hörnfeldt, B. 2005. Vådan av varmare vintrar för sork och lämmel. Miljöforskning 5 (5-6): 40-41. (pdf)
Ecke, F., Christensen, P., Sandström, P. and Hörnfeldt, B. 2006. Identification of landscape elements related to declines of grey-sided vole populations. Landscape Ecology 21: 485-497.
Ecke, F., Magnusson, M. & Hörnfeldt, B. (2013). Spatiotemporal changes in the landscape structure of forests in northern Sweden. Scand. J. For. Res. 28: 651-667.
Ecke, F., Angeler, D. G., Magnusson, M., Khalil, H., and Hörnfeldt, B. 2017. Dampening of population cycles in voles affects small mammal community structure, decreases diversity, and increases prevalence of a zoonotic disease.Ecology and Evolution 7: 5331–5342. doi: 10.1002/ece3.3074
Ecke, F., J. P. Benskin, Å. M. M. Berglund, C. A. de Wit, E. Engström, M. M. Plassmann, I. Rodushkin, D. Sörlin and B. Hörnfeldt (2020) Spatio-temporal variation of metals and organic contaminants in bank voles (Myodes glareolus). Science of The Total Environment: 136353. https://doi.org/10.1016/j.scitotenv.2019.136353.
Hipkiss, T., Gustafsson, J., Eklund, U. and Hörnfeldt, B. 2013. Is the long-term decline of boreal owls in Sweden caused by avoidance of old boxes? J. Raptor Res. 47: 15-20.
Hipkiss, T., Stefansson, O. and Hörnfeldt, B. 2008. Effect of cyclic and declining food supply on great grey owls in boreal Sweden. Can. J. Zool. 86: 1426-1431 (doi: 10.1139/Z08-131) (pdf).
Hörnfeldt, B. 1994. Delayed density dependence as a determinant of vole cycles. Ecology75: 791-806.
Hörnfeldt, B. 1995. Long-term decline in numbers of cyclic voles in northern Sweden. Rapport från Världsnaturfonden WWF Nr. 3: 95, pp. 21-24.
Hörnfeldt, B. 1998. Miljöövervakningen visar på minskande sorkstammar! Fauna och Flora 95: 137-144.
Hörnfeldt, B. 2004. Long-term decline in numbers of cyclic voles in boreal Sweden: analysis and presentation of hypotheses. Oikos 107: 376-392. (pdf)
Hörnfeldt, B., Christensen, P., Sandström, P. & Ecke, F. 2006. Long-term decline and local extinction of Clethrionomys rufocanus in boreal Sweden. Landscape Ecology 21: 1135-1150.
Hörnfeldt, B., Hipkiss, T. & Eklund, U. 2005. Fading out of vole and predator cycles? Proc. Roy. Soc. B 272: 2045-2049. (pdf)
Khalil, H., Ecke, F., Evander, M., Magnusson, M. & Hörnfeldt, B. 2016. Declining ecosystem health and the dilution effect. Scientific Reports, 6, 31314.
Khalil, H., Olsson, G., Magnusson, M., Evander, M., Hörnfeldt, B., and Ecke, F. 2017. Spatial prediction and validation of zoonotic hazard through micro-habitat properties: where does Puumala hantavirus hole – up? Bmc Infectious Diseases 17: 523. doi: 10.1186/s12879-017-2618-z
Khalil H., Ecke F., Evander M., Bucht G., Hörnfeldt B. (2019) Population Dynamics of Bank Voles Predicts Human Puumala Hantavirus Risk. EcoHealth 16 (3): 545–55. https://doi.org/10.1007/s10393-019-01424-4.
Magnusson, M. 2015. Cyclic voles and Puumala hantavirus in a changing boreal landscape. Doktorsavhandling, Sveriges Lantbruksuniversitet, Umeå.
Magnusson, M., Bergsten, A., Ecke, F., Bodin, Ö., Bodin, L. & Hörnfeldt, B. (2013). Predicting grey-sided vole occurrence in northern Sweden at multiple spatial scales. Ecology and Evolution 3: 4365-4376.
Magnusson,_M., Ecke, F., Khalil, H., Olsson, G., Evander, M., Niklasson, & Hörnfeldt, B. 2015. Spatial and temporal variation of hantavirus bank vole infection in managed forest landscapes. Ecosphere 6 (9):163. (http://dx.doi.org/10.1890/ES15-00039.1; published online: 29 September 2015).
Magnusson, M., Hörnfeldt, B. & Ecke, F. 2015. Evidence for different drivers behind long-term decline and depression of density in cyclic voles. Pop. Ecol. 4: 569-580. (dx.doi.org/10.1007/s10144-015-0512-3; published online: 23 September 2015).
Niklasson, B., Nyholm, E., Feinstein, R.E., Samsioe, A. and Hörnfeldt, B. 2006. Diabetes and myocarditis in voles and lemmings at cyclic peak densities – induced by Ljungan virus? Oecologia 150: 1-7.
Läs mer här om annan forskning med anknytning till miljöövervakningen
Projektledare: fr.o.m. 2016-07-01 Frauke Ecke (tidigare Birger Hörnfeldt), Sveriges lantbruksuniversitet